From Surf Wiki (app.surf) — the open knowledge base
Cysteine dioxygenase
Enzyme

Enzyme
| Field | Value |
|---|---|
| Name | Cysteine dioxygenase |
| EC_number | 1.13.11.20 |
| CAS_number | 37256-59-0 |
| image | CDO full structure.png |
| width | 275 |
| caption | Human CDO (drawn from PDB 2IC1) |
| name | Cysteine dioxygenase |
Function
CDO is responsible for the first major step in metabolism of cysteine. CDO oxidizes to cysteine sulfinic acid (which exists predominantly in the anionic sulfinate form in vivo). Overall, CDO catalyzes the addition of dioxygen (O2) to a thiol, producing a sulfinic acid. More specifically, CDO is part of the group of non-heme iron oxygenases that employ oxygen as an electron acceptor. Cysteine sulfinic acid is then metabolized further via two divergent pathways: decarboxylated to hypotaurine by sulfinoalanine decarboxylase and oxidized to taurine by hypotaurine dehydrogenase; or transaminated to a putative 3-sulfinylpyruvate intermediate, which decomposes spontaneously into pyruvate and sulfite. Sulfite can then be oxidized to sulfate by sulfite oxidase. Thus CDO is necessary for hypotaurine/taurine and sulfite/sulfate production. The role of CDO may vary between cell types as it can either be used primarily for taurine or sulfate production or for degradation of cysteine. :[[Image:Cysteine dioxygenase reaction.svg|thumb|left|500px|CDO reaction scheme showing cysteine sulfinic acid formation from cysteine by dioxygen incorporation]]
Structure
CDO is a 22.5 kDa protein that contains 200 amino acid residues and has an isoelectric point (pI) of 5.5. The primary structure is highly conserved between mammalian species, with murine and human CDO differing in only 16 residues. CDO is part of the cupin superfamily, whose members possess a 6-stranded β-barrel in a "jelly-roll" topology. Crystal structures of the protein have been obtained at 1.5 Å resolution (mouse). The active site displays a unique geometry where instead of the typical facial triad of two histidines and one carboxylate side-chain coordinating to an iron (II) species, three histidine ligands are bound to iron. Furthermore, crystal structures show the amino nitrogen and thiolate sulfur of cysteine coordinated to the iron in addition to a single water molecule (see figure).
CDO contains a unique internal cofactor created by intramolecular thioether formation between Cys93 and Tyr157, which is postulated to participate in catalysis. When the protein was first isolated, two bands on agarose gel were observed, corresponding to the cofactor-containing protein and the unlinked "immature" protein, respectively. Crosslinking increases efficiency of CDO ten-fold and is regulated by levels of cysteine, an unusual example of protein cofactor formation mediated by substrate (feedforward activation).
Mechanism
The CDO mechanism is still not well understood, despite active research to elucidate details of the reaction. Overall, the reaction involves addition of O2 to cysteine, which occurs spontaneously without enzyme catalysis. Studies have shown that the cysteinyltyrosine bridge lowers the oxidation potential of tyrosine (commonly an electron donor, as in photosystem II) by ~0.5 V relative to phenol and increases its acidity. The thioether moiety likely plays a structural, redox, or, acid/base role. Other studies have shown that Tyr157 is needed for enzyme function (possibly as a tyrosinyl radical) and is highly conserved across CDO variants. Furthermore, research has shown that cysteamine, a structurally similar molecule to cysteine, enhances cysteine oxidation but is not a substrate.
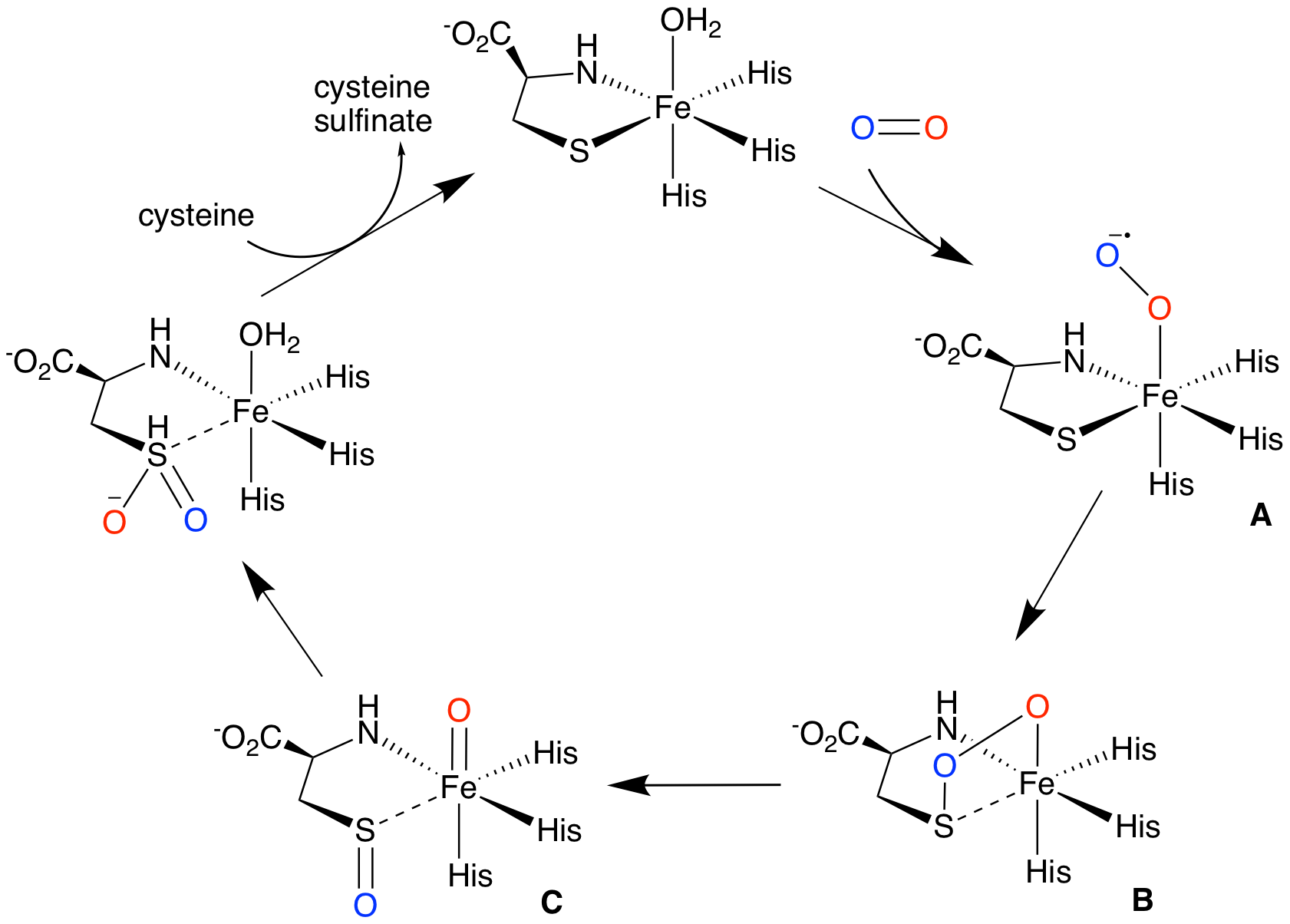
One proposed mechanism, supported by computational and spectroscopic studies, involves O2 binding cis to a thiolate to form reactive iron (III)-superoxo species (A), which then attacks the bound sulfur of cysteine to form a four-membered ring structure (B). Heterolytic O-O bond cleavage then affords a high-valent iron (IV) oxo intermediate (C), which transfers the second oxygen to sulfur.
Regulation
CDO is tightly regulated in the cell to maintain cysteine homeostasis. In particular, CDO responds to changes in dietary cysteine availability and protein intake, maintaining decreased activity with low cysteine levels and increased activity at high levels to prevent cytotoxicity. Studies have shown that CDO can exhibit a dramatic increase in hepatic activity within hours. Unlike many enzymes, it is predominantly regulated at the level of protein turnover rather than transcriptional (mRNA levels). High cysteine levels inhibit ubiquitinylation, which lowers the rate of proteasomal degradation. CDO is also regulated in adipose tissue, where high cysteine levels cause increased hypotaurine/taurine production. Regulation of CDO is also thought to involve both the crosslinked and immature forms of the protein.
Disease Relevance
Because of its relevance to cysteine metabolism, changes in CDO activity may cause disease in humans. Research has found that elevated cysteine can by cytotoxic, neurotoxic, and excitotoxic. Abnormal or deficient CDO activity has been linked to Alzheimer's disease, Parkinson's disease, rheumatoid arthritis, and motor neuron diseases. In these diseases, patients display depressed sulfate levels, elevated fasting cysteine plasma concentrations, and other symptoms consistent with impaired cysteine oxidation. CDO deficiency and subsequent cysteine accumulation in the globus pallidus has been linked to Pantothenate kinase-associated neurodegeneration.
The expression of CDO is altered in cancer cells and methylation of the CDO1 (human cysteine dioxygenase type I) promoter gene was shown to occur in colon, breast, esophageal, lung, bladder, and stomach cancers. Silencing of CDO1 is a critical epigenetic event in breast cancer, leading to downregulation of CDO1 activity. In particular, decreased CDO1 activity causes increased hydrogen sulfide (H2S), which has been connected to various diseases. These results suggest that CDO1 (human cysteine dioxygenase type I) acts as a tumor suppressor gene and may potentially serve as a biomarker for cancer.
References
References
- (May 2009). "Cysteine dioxygenase: a robust system for regulation of cellular cysteine levels". Amino Acids.
- (August 2007). "Cysteine dioxygenase: structure and mechanism". Chemical Communications.
- (June 2011). "Thiol dioxygenases: unique families of cupin proteins". Amino Acids.
- (March 2005). "Heterologous expression, purification, and characterization of recombinant rat cysteine dioxygenase". The Journal of Biological Chemistry.
- (March 1969). "Cystein oxygenase. II. Studies on the mechanism of the reaction with 18oxygen". The Journal of Biological Chemistry.
- (February 1976). "Purification and some properties of rat liver cysteine oxidase (cysteine dioxygenase)". Biochimica et Biophysica Acta (BBA) - Enzymology.
- (February 2007). "An insight into the mechanism of human cysteine dioxygenase. Key roles of the thioether-bonded tyrosine-cysteine cofactor". The Journal of Biological Chemistry.
- (February 2006). "Structure and mechanism of mouse cysteine dioxygenase". Proceedings of the National Academy of Sciences of the United States of America.
- (July 2010). "Spectroscopic and computational characterization of substrate-bound mouse cysteine dioxygenase: nature of the ferrous and ferric cysteine adducts and mechanistic implications". Biochemistry.
- (July 2016). "An iron-oxygen intermediate formed during the catalytic cycle of cysteine dioxygenase". Chemical Communications.
- (January 2017). "Metal-Assisted Oxo Atom Addition to an Fe(III) Thiolate". Journal of the American Chemical Society.
- (March 2011). "Theoretical study on the mechanism of the oxygen activation process in cysteine dioxygenase enzymes". Journal of the American Chemical Society.
- (July 1992). "Genetic factors influencing the outcome of early arthritis--the role of sulphoxidation status". British Journal of Rheumatology.
- (March 1990). "Plasma cysteine and sulphate levels in patients with motor neurone, Parkinson's and Alzheimer's disease". Neuroscience Letters.
- (June 2006). "Mammalian cysteine metabolism: new insights into regulation of cysteine metabolism". The Journal of Nutrition.
- (September 2012). "Cysteine dioxygenase 1 is a tumor suppressor gene silenced by promoter methylation in multiple human cancers". PLOS ONE.
- (June 2013). "Frequent inactivation of cysteine dioxygenase type 1 contributes to survival of breast cancer cells and resistance to anthracyclines". Clinical Cancer Research.
This article was imported from Wikipedia and is available under the Creative Commons Attribution-ShareAlike 4.0 License. Content has been adapted to SurfDoc format. Original contributors can be found on the article history page.
Ask Mako anything about Cysteine dioxygenase — get instant answers, deeper analysis, and related topics.
Research with MakoFree with your Surf account
Create a free account to save articles, ask Mako questions, and organize your research.
Sign up freeThis content may have been generated or modified by AI. CloudSurf Software LLC is not responsible for the accuracy, completeness, or reliability of AI-generated content. Always verify important information from primary sources.
Report